
Fish hearts are S-shaped and four-chambered with, from behind forwards, sinus venosus, atrium, ventricle and either a bulbus or conus leading to the ventral aorta. We have already seen that some airbreathing fishes have a double circulation (the Japanese mudfish, Channa argus, even having a double ventral aorta), but only lungfishes have a morphologically partially divided heart.

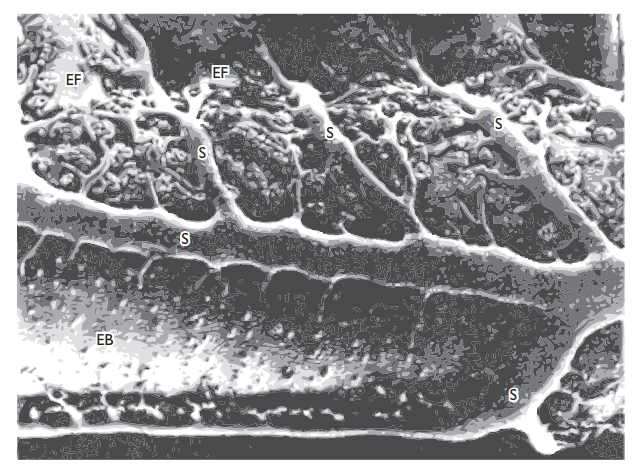
The teleost bulbus is an elastic reservoir passively enlarged by blood driven forwards out of the ventricle, but the equivalent conus (in elasmobranchs, Amia, Lepisosteus and Polypterus) is contractile, contracting in sequence with the rest of the heart. Valves at the junctions between the different regions assure unidirectional flow, and pocket valves are also found along the conus, sometimes in large numbers; Lepisosteus has no less than 72 valves in eight rows. Heart mass in most fishes scales as body mass (as in other vertebrates), but the size of the ventricle differs a good deal, being largest in tunas and icefishes where Q is greatest.
The cardiac cycle consists of systole, when the ventricle is emptied, and diastole when it is refilled; it is accompanied by a progression of electrical events along the heart resulting from the depolarization and repolarization of the cardiac muscle cells. The sum of these cardiac action potentials, which are relatively easy to record in situ with electrodes which need not be in or on the heart itself, are electrocardiograms (ECGs). Fish ECGs are essentially the same as those of mammals: atrial contraction produces the P wave, ventricular contraction the QRS complex, and ventricular relaxation the T wave. But fish heart ECGs can be more complex than in mammals. Contraction of the conus in elasmobranches adds a small B wave, and contraction of the sinus venosus adds a V wave, prominent in the hagfish (Eptatretus) and in the eel.
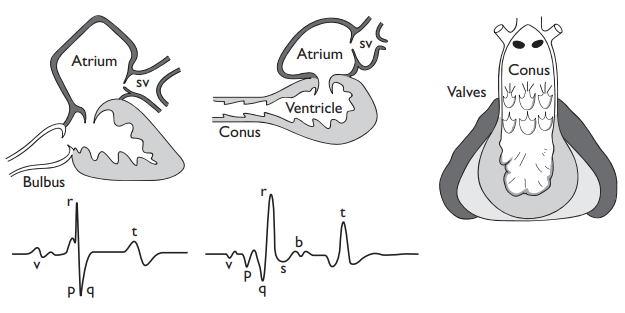
Contractions of the atrium and ventricle have to be coordinated in such a way that there is a delay between atrial and ventricular contraction, and a rapid synchronous contraction of the ventricle. The specially modified conduction pathways (His-Purkinje fibers) found in higher vertebrates seemed to be lacking in fish hearts, and it is only recently that Sedmera et al. (2002) recognized their equivalent in zebra fish.
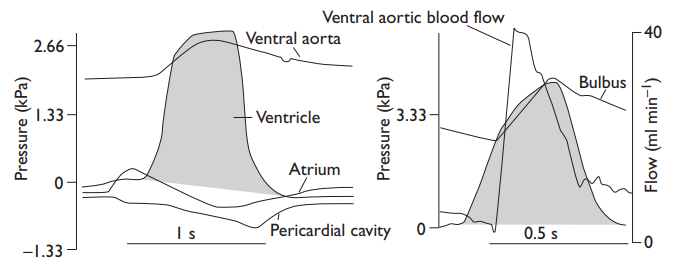
Ventricular contractions naturally lead to cyclic variations in pressure and flow in the ventral aorta from which blood flows through the serial capillary resistances of the gill and systemic capillary beds. Such oscillations are damped in teleosts by the elastic bulbus (constant flow is what is optimal for the gill gas exchanger), but it does not seem that the elasmobranch conus can act in this way, although it has been claimed to do so; conus contraction is too slow to do other than make the pocket valves close together to prevent backflow, and pressures oscillate much more in the ventral aorta than in teleosts.