
The vast majority of living fishes are predatory. Of some 1100 species of extant elasmobranchs, only 13 (1.2%) – the basking shark (Cetorhinus maximus), megamouth shark (Megachasma pelagios), whale shark (Rhincodon typus), manta ray (Manta birostris), and about nine species of devil rays (genus Mobula) – have forsaken the actively predacious habits of their kin and adapted to a more placid “grazing” lifestyle.
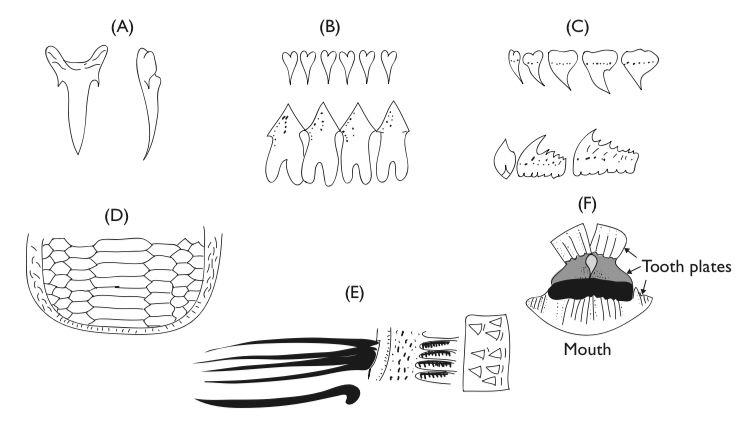
Predatory (piscivorous) fishes typically have large, terminal or subterminal mouths with well developed grasping and biting teeth. Many also have teeth on the tongue and roof of mouth. Among predatory fishes adapted to feeding on hard, shelly prey some jaw or pharyngeal teeth are usually heavy and molariform. Gill rakers, if present, are typically short or blunt. They have a well defined stomach, and typically possess pyloric caecae. The intestine is relatively short. They may be strong swimmers or sit-and-wait ambushers. And, if the latter, they may possess camouflage, lures and the like.
Some predatory fishes feed on the sea’s abundant gelatinous animals such as ctenophores, jellyfishes, and salps. Correlated with a gelatinous diet are enlarged digestive tracts, exceptionally large stomachs, and extremely long intestines. Fish feeding on gelatinous animals also may have pharyngeal or esophageal modifications, presumably to prevent regurgitation (Fänge and Grove, 1979). Mola mola has three rows of recurved pharyngeal teeth. In the stromateoid fishes, there are esophageal sacs with denticulate papillae. Genicanthus personatus, the masked angelfish, includes hydromedusae and siphonophores in its diet and has finger-like esophageal papillae that point posteriorly. The esophagus of chum salmon is strongly muscular with a well define sphincter.
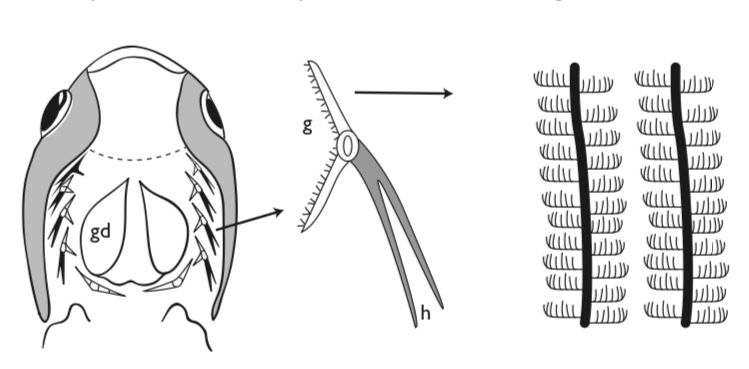
Another major feeding guild consists of plankton filterers. A successful plankton filterer needs to be able to separate plankton from the water. Typically this occurs through mechanical filtration by means of gill rakers, but, as noted in “Food choices, food and development”, aerosol filtering is employed as well. Some filter-feeders, especially those in which phytoplankton is an important source of food, have exceedingly long guts. The gut in the menhaden is ten times the length of that in the herring and has 400 pyloric caecae compared with 20 in the herring, reflecting the greater difficulty of digesting plant food. Filter-feeding appears during ontogeny; it is not found in young larvae, which are usually microzooplankton particle-feeders. In herring and menhaden, in the paddle fish Polyodon and in certain cichlid species filtering appears as the gill rakers develop. Most of the clupeoids must reach a length of 80–100 mm before starting to filter.
There are a few large fishes that are able to filter sufficient water to be able to filter-feed on zooplankton when this is abundant. In freshwater, the paddle fish (Polyodon) of the Mississippi Basin filters cladocerans, cruising around with its lower jaw dropped almost to 90. In the oceans, the huge whale sharks (Rhincodon), basking sharks (Cetorhinus), and manta and devil rays (Mobula) all filter-feed. In whale sharks and manta rays, the filter is spongy tissue formed from modified denticles, while in basking sharks, there are long gill rakers (again modified denticles;) on gill arch. Whale sharks sometimes filter in the vertical position, pushing their heads slowly out of the water, allowing the pharynx to drain, and then subsiding slowly again below the surface. Basking sharks feed on copepods and live in temperate waters, where zooplankton varies widely in abundance during the year.
While early calculations based on assumed zooplankton calorific value and abundance on the one hand, and estimates of the energy expenditure of the shark when swimming with wide open mouth while filtering on the other, suggested that these enormous fish are actually balanced on the knife edge of using more energy capturing their food than they can obtain from it. Several of the assumptions behind these calculations were recently shown by David Sims (1999, 2000; Sims et al., 2003) to be faulty, resulting in an overestimation (by about three times) of the energy basking sharks require to swim. Also, contrary to earlier reports, basking sharks (although not all of them) do not shed their gill rakers and hibernate in the winter, but instead they feed on deeper-dwelling populations of mesopelagic zooplankton that do not experience such dramatic seasonal variations in abundance. However, at least some sharks do shed their gill rakers; one stranded at Polperro in Cornwall in November had the rakers covered with epithelium and shorter than those of feeding adults.
Particle-feeding or picking zooplankton is seen also in some rather unexpected fishes. The deep-sea tripod fishes pick copepods from the benthopelagic plankton, as they sit perched on their fins facing into the current. Curiously, the spur dog (Squalus) may also feed in this way on the planktonic ctenophore Pleurobrachia, which it sometimes takes in such amounts as to fill its gut, and it has recently been suggested (from their parasite communities) that eels of all sizes in rivers and lakes in England feed regularly on planktonic copepods.
Many detritivores are bottom vacuumers/suckers that typically possess small, undershot, inferior mouths equipped with suctorial lips and barbels, sensory pits, etc. positioned to help detect buried prey. Other detritivores feed on phytoplankton and mud particles, triturated by the same kind of gizzard as in mullets (Mugilidae). Although mullets typically browse on the algal film over mud (leaving a characteristic series of depressions where they have gulped in surface mud), they can also feed on planktonic algae at the surface–air interface using the gill rakers as a sieve. Detritivores may have either a one or two-part stomach. Those with two-part stomachs have short intestines and those with a one-part stomach, a long intestine.
Herbivorous fishes Herbivorous (phytophagous) fishes generally have small, often inferior, mouths equipped with rasping or nipping teeth. Pharyngeal teeth may be present. Stomachs are generally absent, but if present are thin walled and elastic. Because plant material is usually difficult to digest they may use cellulase and other enzymes produced by gut microflora (Jobling, 1995), hind gut fermentation by bacteria and protozoans of carbohydrates to short-chain fatty acids that can be directly absorbed with intervention of bile salts, or mechanical processing (trituration) by a muscular stomach/gizzard or mastication by pharyngeal teeth.
Overall, herbivorous fishes are very much in the minority, and are least common in the sea and in temperate freshwaters. Only some 5% of all fish families (18 marine and at least 20 freshwater) contain herbivores, although herbivorous individuals may be among the most numerous in any fish community (Horn, 1989). Perhaps this is because although more energy has to be expended by herbivores than by carnivores in obtaining their food, herbivrous food resources tend to be quite abundant. Certainly, compared with carnivores, herbivores have to spend a good deal more of their lives feeding. Some phytoplankton feeders filter continuously, while a parrot fish spent 8 h day –1, and Haplochromis and Tilapia 14 h day –1, this in comparison with 1–3 h day–1 for salmonids. Whatever the diet, all fish have a high protein requirement (around 50% of dry weight of the diet). How do herbivores satisfy this requirement? As detritivores and herbivores they ingest large amounts of attached protein-rich microorganisms, even if they do not eat protein-rich algae.
Most marine herbivores are found around the coral reefs of the tropics, where surgeon fishes (Acanthuridae) crop algae growing over corals and on sandy patches near the reef, while parrot fishes (Scaridae), rabbit fishes (Siganidae), and damsel fishes (Pomacentridae) browse algae scraped off coral surfaces. Some temperate shore fishes, such as blennies and gobies, feed largelyon seaweeds, and the much prized temperate freshwater ayu (Plecoglossus) of Japanese rivers, feeds solely on moss scraped from its territory of mossy stones. Ayu are an expensive delicacy since they are caught in a curious and somewhat inefficient way; fishermen place a small ayu on their line, below which is a series of unbaited hooks. Attracted by this decoy, another ayu is foul-hooked as it comes out to defend its feeding territory.
Herbivory is, however, much more common in tropical freshwaters, where seasonal flooding inundates forests and plains. Here fishes such as the grass carp (Ctenopharyngodon) can feed on grasses, and decaying vegetation, while fruits, flowers, and seeds that fall into the water are seized by fishes such as the cyprinid Puntius or fruit and nut eating characins, including the tambaqui (Colossoma macropomum) which can attain a weight of 30 kg, and species of piranhas (Serrasalmus). As in phytoplankton feeders, filter-feeders, and mammalian herbivores, herbivorous fish have longer guts with greater surface area for absorption than carnivores or omnivores. As a rough rule, the ratio of gutlength to body length is greater than 3 in herbivorous fishes, from 1 to 3 in omnivores and less than 1 in carnivores.
Fish have also evolved adaptations that permit them to feed on the fins, scales, mucus, blood (lampreys, candiru (Vandellia cirrhosa), male angler fish), even the eyes (Haplochromis compressiceps) of other organisms. Such species typically possess highly specialized, often bizarre morphological and behavioral adaptations. Cleaner fish that feed on ectoparasites also provide interesting examples of symbiotic relationships between the small cleaner and the usually much larger host species as described .
Food and feeding presents some of the most interesting applications of cur- rent ecological theory and modern technology in the study of the lives of fishes. How fishes acquire their food, what they eat, and how that food is uti- lized are all fundamental questions facing investigators. Traditional studies, often based on stomach contents, are now enhanced by computer modeling, electromyographic recording, and digital photographic techniques that allow for real-time, in-vivo data collection on feeding habits.