
Cephalopoda (Gr. kephal¯e, head, + pous, podos, foot) include squids, octopuses, nautiluses, devilfish, and cuttlefish. All are marine, and all are active predators. Their modified foot is concentrated in the head region. It takes the form of a funnel for expelling water from the mantle cavity, and the anterior margin is drawn out into a circle or crown of arms or tentacles.
Cephalopods range upward in size from 2 or 3 cm. The common squid of markets, Loligo, is about 30 cm long. Giant squids, Architeuthis, at up to almost 60 ft in length and weighing nearly a ton, are the largest invertebrates known.
Fossil records of cephalopods go back to Cambrian times. The earliest shells were straight cones; others were curved or coiled, culminating in the coiled shell similar to that of the modern Nautilus, the only remaining member of the once flourishing nautiloids. Cephalopods without shells or with internal shells (such as octopuses and squids) apparently evolved from some early straight-shelled ancestor. Many ammonoids, which are extinct, had quite elaborate shells.
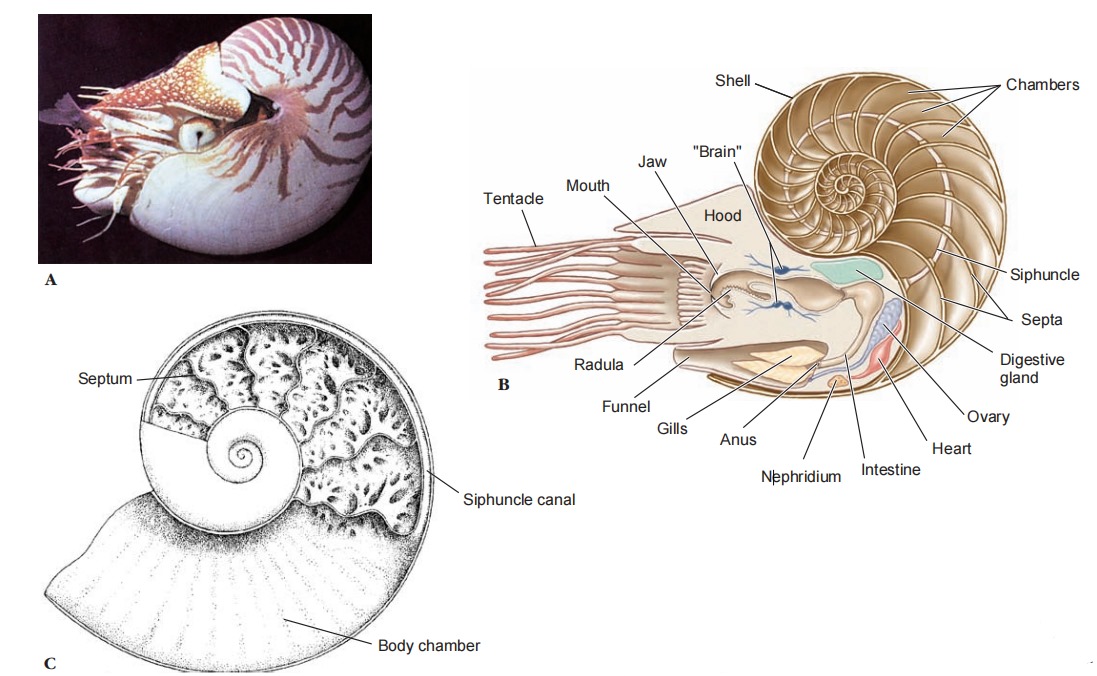
and diagram of body structure. C, Longitudinal section through shell of an ammonoid.
The natural history of some cephalopods is fairly well known. They are marine animals and appear sensitive to the degree of salinity. Few are found in the Baltic Sea, where the water has a low salt content. Cephalopods are found at various depths. Octopuses are often seen in the intertidal zone, lurking among rocks and crevices, but occasionally they are found at great depths. The more active squids are rarely found in very shallow water, and some have been taken at depths of 5000 m. Nautilus is usually found near the bottom in water 50 to 560 m deep, near islands in the southwestern Pacific.
The enormous giant squid, Architeuthis, is very poorly known because no one has ever studied a living specimen. The anatomy has been described from stranded animals, from those captured in nets of fishermen, and from specimens found in the stomach of sperm whales. The mantle length is 5 to 6 m, and the head is up to 1 m. They have the largest eyes in the animal kingdom: up to 25 cm (10 in) in diameter. They apparently eat fish and other squids, and they are an important food item for sperm whales. They are thought to live on or near the sea bottom at a depth of 1000 m, but some have been observed swimming at the surface.
Form and Function
Shell Although early nautiloid and ammonoid shells were heavy, they were made buoyant by a series of gas chambers, as is that of Nautilus, enabling the animal to maintain neutral buoyancy. The shell of Nautilus, although coiled, is quite different from that of a gastropod. The shell is divided by transverse septa into internal chambers, only the last inhabited by the living animal. As it grows, it moves forward, secreting behind its body a new septum. The chambers are connected by a cord of living tissue called a siphuncle, which extends from the visceral mass. Cuttlefishes also have a small, curved shell, but it is entirely enclosed by the mantle. In squids most of the shell has disappeared, leaving only a thin, proteinaceous strip called a pen, which is enclosed by the mantle. In Octopus (Gr. oktos, eight, pous, podos, foot) the shell has disappeared entirely.

After a member of genus Nautilus secretes a new septum, the new chamber is filled with fluid similar in ionic composition to that of Nautilus’ s blood (and of seawater). Fluid removal involves active secretion of ions into tiny intercellular spaces in the siphuncular epithelium, so that a very high local osmotic pressure is produced, and water is drawn out of the chamber by osmosis.
The gas in the chamber is just the respiratory gas from the siphuncle tissue that diffuses into the chamber as fluid is removed. Thus gas pressure in the chamber is 1 atmosphere or less because it is in equilibrium with gases dissolved in the seawater surrounding the Nautilus, which are in turn in equilibrium with air at the surface of the sea, despite the fact that the Nautilus may be swimming at 400 m beneath the surface. That the shell can withstand implosion by the surrounding 41 atmospheres (about 600 pounds per square inch), and that the siphuncle can remove water against this pressure are marvelous feats of natural engineering!
Locomotion Cephalopods swim by forcefully expelling water from the mantle cavity through a ventral funnel (or siphon)—a sort of jet propulsion. The funnel is mobile and can be pointed forward or backward to control direction; the force of water expulsion controls speed. Squids and cuttlefishes are excellent swimmers. The squid body is streamlined and built for speed. Cuttle-fishes swim more slowly. The lateral fins of squids and cuttle-fishes serve as stabilizers, but they are held close to the body for rapid swimming. Nautilus is active at night; its gas-filled chambers keep the shell upright. Although not as fast as squids, it moves surprisingly well.
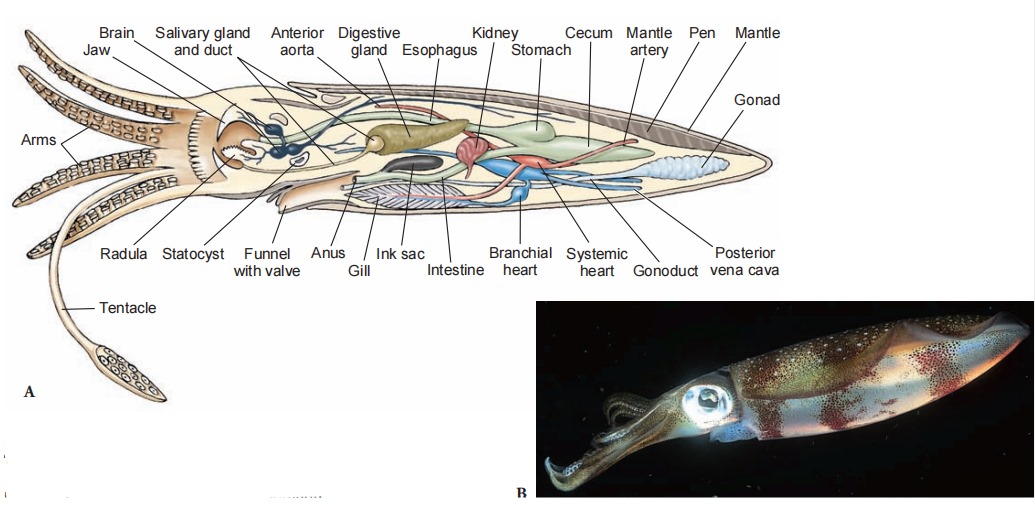
Octopus has a rather globular body and no fins. An octopus can swim backward by spurting jets of water from its funnel, but it is better adapted to crawling over rocks and coral, using suction discs on its arms to pull or to anchor itself. Some deep-water octopods have the arms webbed like an umbrella and swim in a medusa-like fashion.
Internal Features
The active habits of cephalopods are reflected in their internal anatomy, particularly their respiratory, circulatory, and nervous systems.
Respiration and Circulation.
Except for nautiloids, cephalopods have one pair of gills. Because ciliary propulsion would not circulate enough water for their high oxygen requirements, there are no cilia on the gills. Instead, radial muscles in the mantle wall compress the wall and enlarge the mantle cavity, drawing water inside. Strong circular muscles contract and expel water forcibly through the funnel. A system of one-way valves prevents water from being taken in through the funnel or expelled around the mantle margin.
Likewise, the open circulatory system of ancestral molluscs would be inadequate for cephalopods. Their circulatory system has evolved into a closed network of vessels, and capillaries conduct blood through the gill filaments. Furthermore, the molluscan plan of circulation places the entire systemic circulation before the blood reaches the gills (in contrast to vertebrates, in which the blood leaves the heart and goes directly to the gills or lungs). This functional problem was solved by the development of accessory or branchial hearts at the base of each gill to increase the pressure of blood going through the capillaries there.
Nervous and Sensory Systems.
Nervous and sensory systems are more elaborate in cephalopods than in other molluscs. The brain, the largest in any invertebrate, consists of several lobes with millions of nerve cells. Squids have giant nerve fibers (among the largest known in the animal kingdom), which are activated when the animal is alarmed and initiate maximal contractions of the mantle muscles for a speedy escape.
Squid nerves played an important role in early biophysical studies. Our current understanding of transmission of action potentials along and between nerve fibers is based primarily on work performed using the giant nerve fi bers of squids, Loligo spp. A. Hodgkin and A. Huxley received the Nobel Prize in Physiology or Medicine, 1963, for their achievements in this fi eld. Sense organs are well developed. Except for Nautilus,which has relatively simple eyes, cephalopods have highly complex eyes with cornea, lens, chambers, and retina.Orientation of the eyes is controlled by the statocysts, which are larger and more complex than in other molluscs. The eyes are held in a constant relation to gravity, so that the slit-shaped pupils are always in a horizontal position.
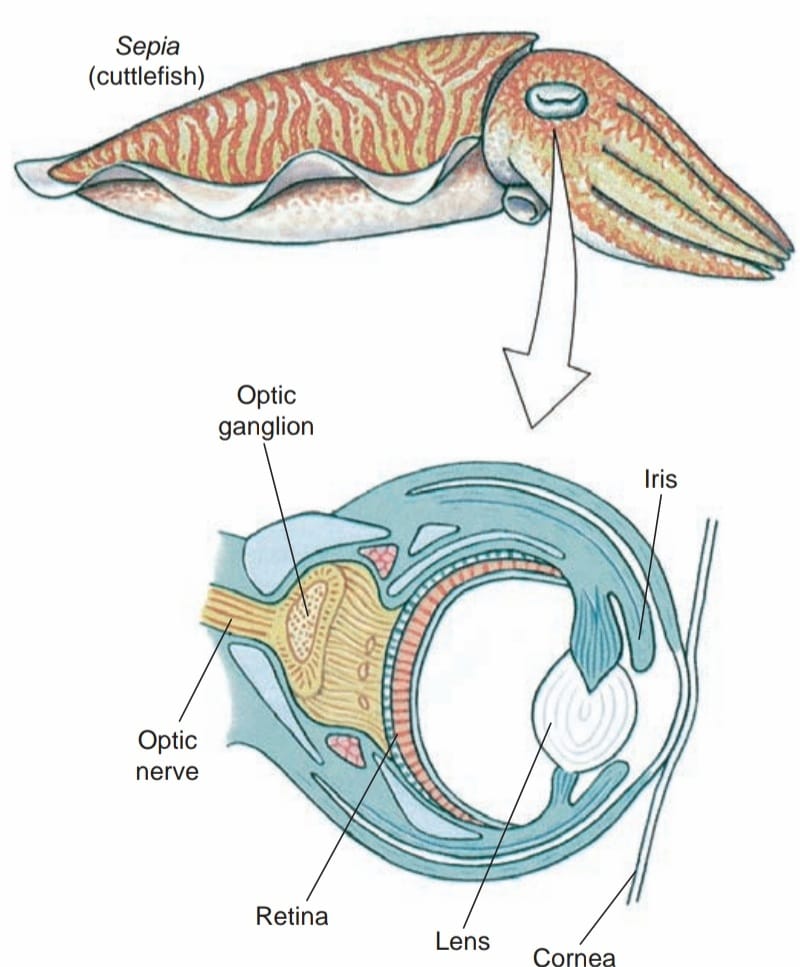
Most cephalopods are apparently colorblind, but their eyesight is excellent and their visual acuity underwater far surpasses our own. They can also be taught to discriminate between shapes for example, a square and a rectangle and to remember such a discrimination for a considerable time. Experimenters find it easy to modify octopod behavior patterns by devices of reward and punishment. Octopods are capable of observational learning; when one octopus observes another being rewarded by making a correct choice, the observer learns which choice is rewarded and consistently makes the same selection when given the opportunity.
When similar structures that are not inherited from a common ancestor evolve in different ways in unrelated animals, we call it convergence, or convergent evolution. For many years cephalopod eyes and vertebrate eyes have been cited as a marvelous example of convergent evolution. Cephalopod and vertebrate eyes are similar in many details of structure but differ in development. Compound eyes of arthropods, differing in both structure and development, were viewed as examples of other, independently derived eyes in animals.
Now we recognize that all triploblastic animals with eyes, even those with the most simple eyespots, such as platyhelminths, share at least two conserved genes: that for rhodopsin, a visual pigment, and Pax 6, now sometimes called the “master control gene for eye morphogenesis.” Once these two genes originated, natural selection eventually produced the specialized organs of vertebrates, molluscs, and arthropods.
Octopods use their arms for tactile exploration and can dis criminate between textures by feel but apparently not between shapes. Their arms are well supplied with both tactile and chemoreceptor cells. Cephalopods seem to lack a sense of hearing.
Communication
Little is known of social behavior of nautiloids or deep-water cephalopods, but inshore and littoral forms such as Sepia, Sepioteuthis, Loligo, and Octopus have been studied extensively. Although their tactile sense is well developed and they have some chemical sensitivity, visual signals are the pre dominant means of communication. These signals consist of a host of movements of the arms, fins, and body, as well as many color changes. Movements may range from minor body motions to exaggerated spreading, curling, raising, or lowering of some or all of the arms. Color changes are effected by chromatophores, cells in the skin that contain pigment granules.
Tiny muscle cells surround each elastic chromatophore, whose contractions pull the cell boundary of the chromatophore outward, causing it to expand greatly. As the cell expands, the pigment becomes dispersed, changing the color pattern of the animal. When the muscles relax, chromatophores return to their original size, and pigment becomes concentrated again. By means of the chromatophores, which are under nervous and probably hormonal control, an elaborate system of changes in color and pattern is possible, including general darkening or lightening; flushes of pink, yellow, or lavender; and formation of bars, stripes, spots, or irregular blotches. These colors may be used variously as danger signals, as protective coloring, in courtship rituals, and probably in other ways.
By assuming different color patterns of different parts of the body, a squid can transmit three or four different messages simultaneously to different individuals and in different directions, and it can instantaneously change any or all of the messages. Probably no other system of communication in invertebrates can convey so much information so rapidly.
Deep-water cephalopods may have to depend more on chemical or tactile senses than their littoral or surface cousins, but they also produce their own type of visual signals, for they have evolved many elaborate luminescent organs. Most cephalopods other than nautiloids have another protective device. An ink sac that empties into the rectum contains an ink gland that secretes sepia, a dark fluid containing the pigment melanin, into the sac. When the animal is alarmed, it releases a cloud of ink, which may hang in the water as a blob or be contorted by water currents. The animal quickly departs from the scene, leaving the ink as a decoy to the predator.
Reproduction: Sexes are separate in cephalopods. Spermatozoa are encased in spermatophores and stored in a sac that opens into the mantle cavity. One arm of adult males is modified as an intromittent organ, called a hectocotylus, used to pluck a spermatophore from his own mantle cavity and insert it into the mantle cavity of a female near the oviduct opening. Before copulation males often undergo color displays, apparently directed against rival males. Eggs are fertilized as they leave the oviduct and are then usually attached to stones or other objects. Some octopods tend their eggs. Females of Argonauta, the paper nautilus, secrete a fluted “shell,” or capsule, in which eggs develop.
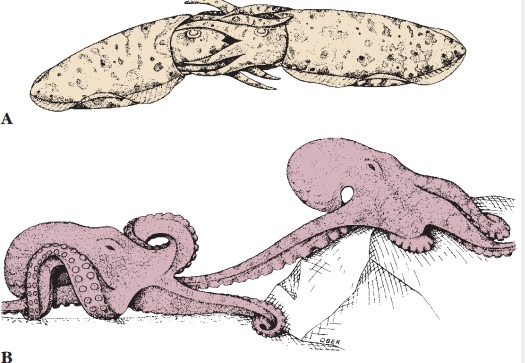
fertilize her eggs. Octopuses often tend their eggs during development.
The large yolky eggs undergo meroblastic cleavage. During embryonic development, the head and foot become indistinguishable. The ring around the mouth, which bears the arms, or tentacles, may be derived from the anterior part of the foot. Juveniles hatch from eggs; no free-swimming larva exists in cephalopods.
Useful External Links