
It is not at all clear why some species have evolved a strategy of producing many small eggs, and others the opposite strategy of fewer, larger ones. The relationships between these characters and the implication for the species can be summarized (Blaxter, 1988) as follow:
- Fecundity and egg size are inversely related when comparisons are made between species. In batch spawners, egg size tends to decrease as the spawning season progresses.
- Fecundity increases with age and size of the female within a species
- Large eggs take longer to develop than small eggs, when interspecific comparisons are made.
- Large eggs produce larger larvae at hatching with a longer period of feeding on yolk reserves.
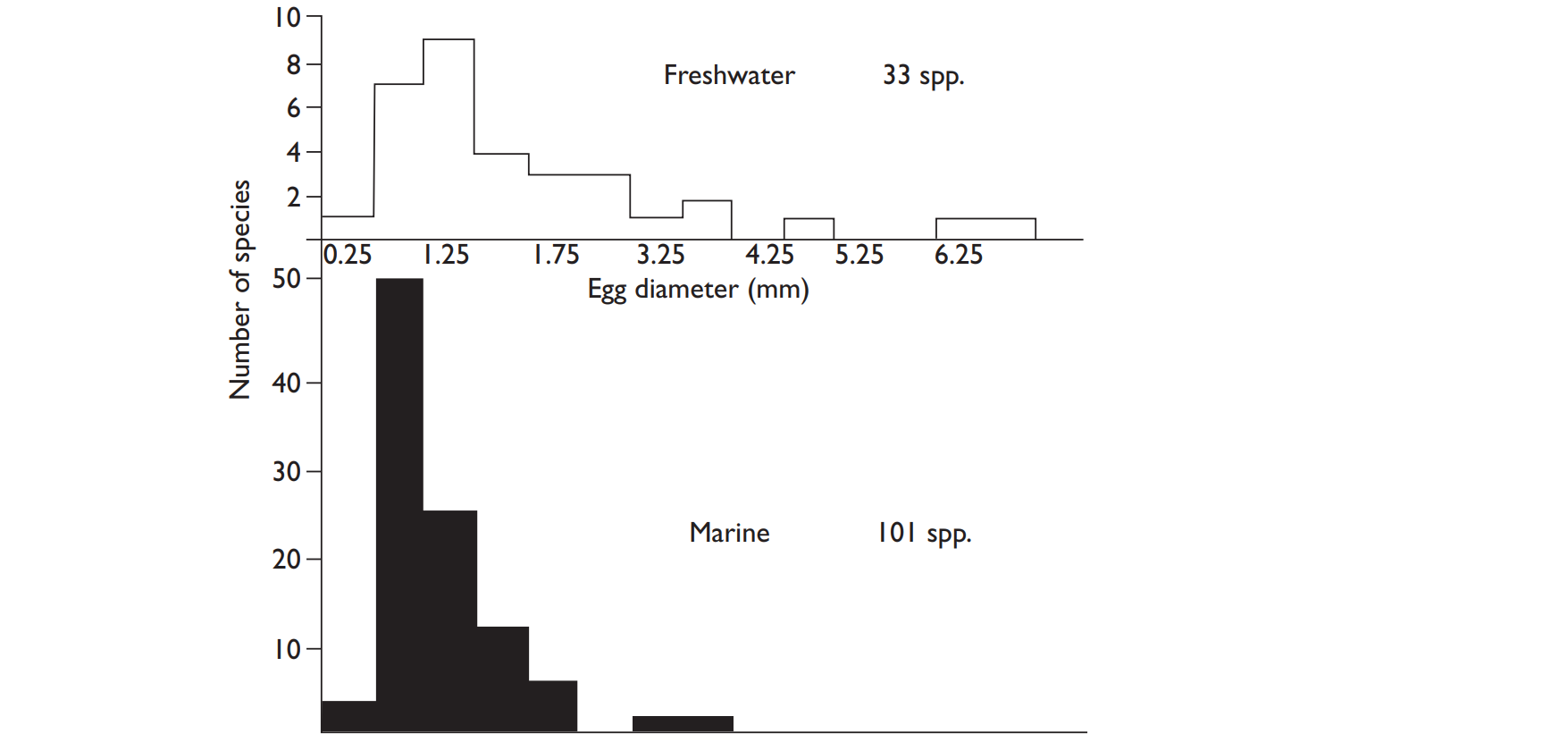
- Fecundity tends to be high in marine fish that release their eggs into open water; it is lower in freshwater species and those species that provide parental
Winemiller and Rose (1992) note that the life history strategies of fishes fall within a trilateral continuum, the endpoints of which comprise three basic strategies: opportunistic spawners such as anchovies, silversides, and most cyprinodontiformes, with high reproductive effort, early maturation, and small body size, low per batch fecundity but multiple spawning bouts per season, little parental investment, and short generation times; periodic spawners, such as salmon, most clupeids and other marine species with moderate reproductive effort, delayed maturity, and large body size contributing to high batch fecundity, little parental investment, and long generation times; and equilibrium spawners, such as cichlids, centrarchids, syngnathids, nest-building coral reef fishes, and many catfishes with low reproductive effort, variable body size and age at maturity producing low batch fecundity but with high parental investment, and moderate to long generation times.
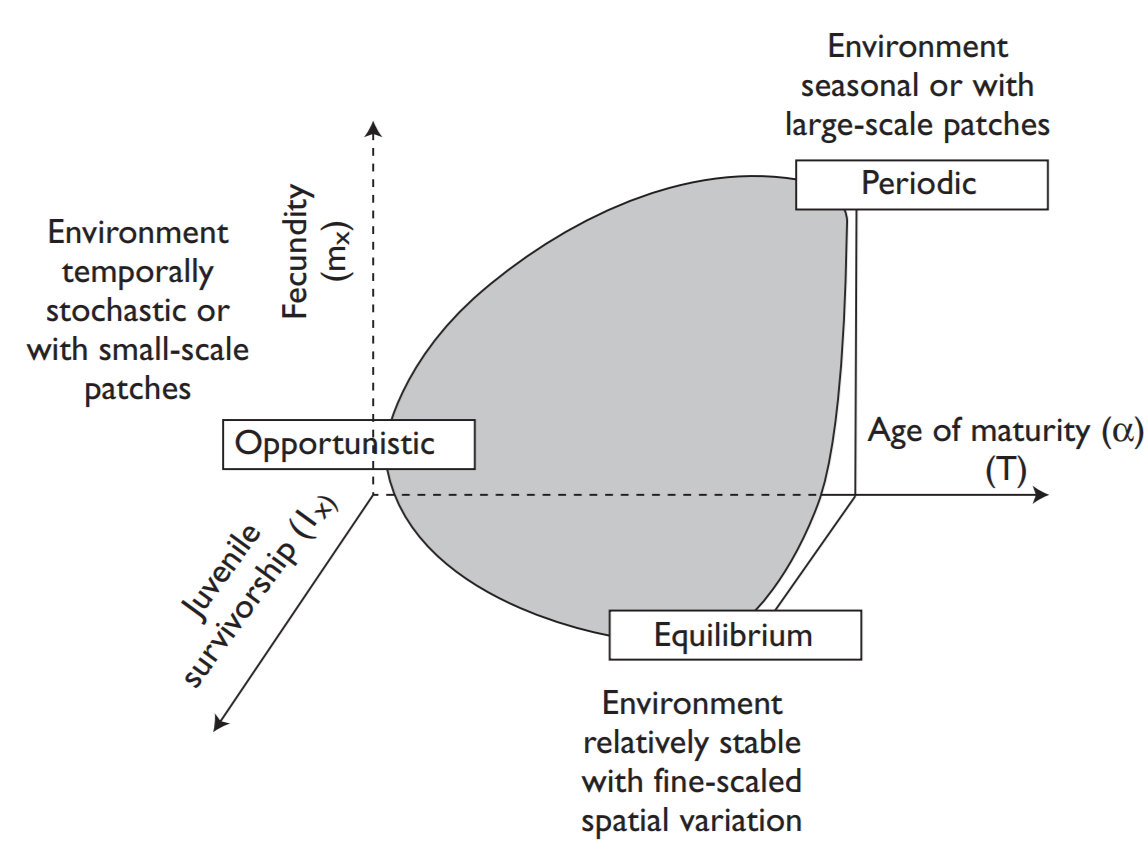
While the three apices of the triangular model correspond to extremes, there are many intermediate examples representing trade-offs between the different factors that determine overall reproductive success in many environments. The second and third categories correspond respectively more-or-less to the “r” and “K” selective strategies applicable to many phyla that have been discussed by MacArthur and Wilson (1967) and other theoretical ecologists. A K strategy is appropriate in stable crowded environments where a low fecundity (large egg) and long developmental period may be favored. In less stable, uncrowded environments, where there are chances for maximal population growth, the r-strategy is optimal with high fecundity (small egg) and a short developmental period to exploit any opportunities for expansion.