Members of phylum Acanthocephala (a-kan’ -tho-sef ‘-a-la) (Gr.akantha, spine or thorn, + kephale¯, head) are commonly called “spiny-headed worms.” The phylum derives its name from one of its most distinctive features, a cylindrical, invaginable proboscis bearing rows of recurved spines, by which it attaches itself to the intestine of its host . The phylum is cosmopolitan, and more than 1100 species are known, most of which parasitize fish, birds, and mammals. All acanthocephalans are endo-parasitic, living as adults in the intestine of vertebrates. Larvae of spiny-headed worms develop in arthropods, either crustaceansor insects, depending on the species.
Various species range in size from less than 2 mm to more than 1 m in length. Females of a species are usually larger than males. Their body is usually bilaterally flattened, with numerous transverse wrinkles. Worms are typically cream color but may absorb yellow or brown pigments from the intestinal contents.
In life the body is somewhat flattened, although students may see turgid and cylindrical specimens that were treated with tap water before fixation . The body wall is syncytial, and its surface is covered by minute crypts 4 to 6 µ m deep, which greatly increase the surface area of the tegument. About 80% of the thickness of the tegument is the radial fiber zone, which contains a lacunar system of ramifying fluid-filled canals.
Exchange of gases, nutrients, and wastes occurs primarily across the body wall by diffusion. Within the body, diffusion is facilitated by the lacunar system. Curiously, body-wall muscles are tubelike and fi lled with fluid. Tubes in the muscles are continuous with the lacunar system; therefore, circulation of lacunar fluid may well bring nutrients to and remove wastes from the muscles. There is no heart or other circulatory system, and contraction of the muscles moves lacunar fluid through the canals and muscles. Thus, the lacunar fluid, which also permeates most tissues of the body, appears to serve as an unusual circulatory system in these animals. Both longitudinal and circular body-wall muscles are present.
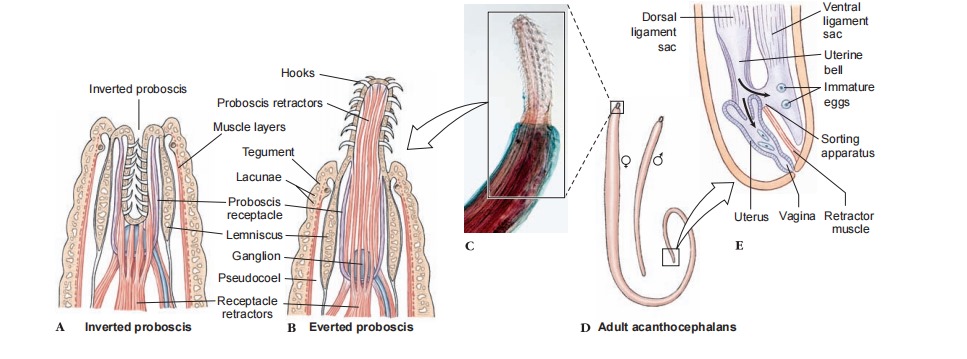
Their proboscis, which bears rows of recurved hooks, is attached to the neck region and can be inverted into a proboscis receptacle by retractor muscles. Attached to the neck region (but not within the proboscis) are two elongated hydraulic sacs (lemnisci) that may serve as reservoirs of lacunar fluid from the proboscis when that organ is invaginated or aid in gas exchange between the body and the proboscis; their exact function remains unknown, however. There is no respiratory system. When present, the excretory system consists of a pair of protonephridia with flame cells. These unite to form a common tube opening into the sperm duct or uterus.
Their nervous system has a central ganglion within the proboscis receptacle and nerves to the proboscis and body. There are sensory endings on the proboscis and genital bursa. However, like many obligate endoparasites, both the nervous system and sense organs of these animals are greatly reduced. Acanthocephalans have no digestive tract, and they must absorb all nutrients through their tegument. They can absorb various molecules by specific membrane-transport mechanisms, and other substances can cross the tegumental membrane by pinocytosis.
Their tegument bears some enzymes, such as peptidases, which can cleave several dipeptides, and the amino acids are then absorbed by the worm. Like cestodes, acanthocephalans require host dietary carbohydrate, but their mechanism for absorption of glucose is different. As glucose is absorbed, it is rapidly phosphorylated and compartmentalized, so that a metabolic “sink” is created into which glucose from the surrounding medium can flow. Glucose diffuses down the concentration gradient into the worm because it is constantly removed as soon as it enters.
Acanthocephalans are dioecious. Males have a pair of testes, each with a vas deferens, and a common ejaculatory duct that ends in a small penis. During copulation sperm are ejected into the vagina, travel up the genital duct, and escape into the pseudocoel of the female.
In females the ovarian tissue in the ligament sac breaks into ovarian balls that are released from the genital ligaments, or ligament sacs, and float free in the pseudocoel. One of the ligament sacs leads to a funnel-shaped uterine bell that receives the developing shelled embryos and passes them to the uterus . An interesting and unique selective apparatus operates here. Fully developed embryos are slightly longer than immature ones, and they are actively selected and passed into the uterus, while immature eggs are rejected and retained for further maturation. The shelled embryos, which are discharged in feces of their vertebrate host, do not hatch until eaten by an intermediate host.
No species is normally a parasite of humans, although species that usually occur in other hosts infect humans occasionally. Macracanthorhynchus hirudinaceus (Gr. makros, long, large, akantha, spine, thorn, rhynchos, beak) occurs throughout the world in the small intestine of pigs and sometimes in other mammals. For M. hirudinaceus the intermediate host is any of several species of soil-inhabiting beetle larvae, especially scarabeids. Grubs of the June beetle (Phyllophaga) are frequent hosts.
Here the larva (acanthor) burrows through the intestine and develops into a juvenile (cystacanth) in the insect’s hemocoel. Pigs become infected by eating the grubs. Acanthocephalans penetrate the intestinal wall with their spiny proboscis to attach to the host. In many cases there is remarkably little inflammation, but in some species the inflammatory response of the host is intense. Infection with these worms can cause great pain, particularly if the gut wall is completely perforated. Multiple infections may do considerable damage to a pig’s intestine, and perforations can occur.
Useful External Links