Nucleus
As in other eukaryotes, the nucleus is a membrane-bound structure whose interior communicates with the cytoplasm by small pores. Within the nucleus the genetic material (DNA) is borne on chromosomes. Except during cell division, chromosomes are not usually condensed in a form that can be distinguished, although during fixation of the cells for light microscopy, chromosomal material (chromatin) often clumps together irregularly, leaving some areas within the nucleus relatively clear. This appearance is described as vesicular and is characteristic of many protozoan nuclei. Condensations of chromatin may be distributed around the periphery of the nucleus or internally in distinct patterns. In most dinoflagellates chromosomes are visible through interphase as they would appear during prophase of mitosis.
Also within the nucleus, one or more nucleoli are often present. Characters such as the persistence of nucleoli during mitosis are useful in identifying protozoan clades. Macronuclei of ciliates are described as compact or condensed because the chromatin material is more finely dispersed and clear areas cannot be observed with the light microscope.

Mitochondria
A mitochondrion is an organelle used in energy production where oxygen serves as the terminal electron acceptor. It contains DNA. Cristae, the internal membranes of a mitochondrion, are of variable form, being flat, tubular, discoid, or branched (ramifying). The form of cristae is considered a homologous character and, in conjunction with other morphological features, is used to describe protozoan clades. In cells without mitochondria, hydrogenosomes may be present. Hydrogenosomes function in the absence of oxygen and are assumed to have evolved from mitochondria. Kinetoplasts are also assumed to be mitochondrial derivatives, but they work in association with a kinetosome, an organelle at the base of a flagellum.
Golgi Apparatus
The Golgi apparatus is part of the secretory system of the endoplasmic reticulum. Golgi bodies are also called dictyosomes in protozoan literature. Parabasal bodies are similar structures with potentially similar functions.
Plastids
Plastids are organelles containing a variety of photosynthetic pigments. The original addition of a plastid to eukaryotic cells occurred when a cyanobacterium was engulfed and not digested. Chloroplasts contain different versions of chlorophylls (a, b, or c), but other kinds of plastids contain other pigments. For example, red algal plastids contain phycobilins. Particular pigments shared among unicellular eukaryotes may indicate shared ancestry, but plastids could also have been gained by secondary endosymbiosis.

Extrusomes
This general term refers to membrane-bound organelles in protozoans that are used to extrude something from the cell. The wide variety of structures extruded suggests that not all extrusomes are homologous. The ciliate trichocyst is an extrusome.
Nutrition
Holozoic nutrition implies phagocytosis, in which an infolding or invagination of the cell membrane surrounds a food particle. As the invagination extends farther into the cell, it is pinched off at the surface. The food particle thus is contained in an intracellular, membrane bound vesicle, a food vacuole or phagosome. Lysosomes, small vesicles containing digestive enzymes, fuse with the phagosome and pour their contents into it, where digestion begins. As digested products are absorbed across the vacuolar membrane, the phagosome becomes smaller. Any undigestible material may be released to the outside by exocytosis, the vacuole again fusing with the cell-surface membrane. In most ciliates, many flagellates, and many apicomplexans, the site of phagocytosis is a definite mouth structure, the cytostome. In amebas,phagocytosis can occur at almost any point by envelopment of a particle with pseudopodia. Particles must be ingested through the opening of the test, or shell, in amebas that have tests. Flagellates may form a temporary cytostome, usually in a characteristic position, or they may have a permanent cytostome with specialized structure. Many ciliates have a characteristic structure for expulsion of waste matter, the cytopyge or cytoproct, found in a characteristic location. In some, the cytopyge also serves as the site for expulsion of the contents of the contractile vacuole.
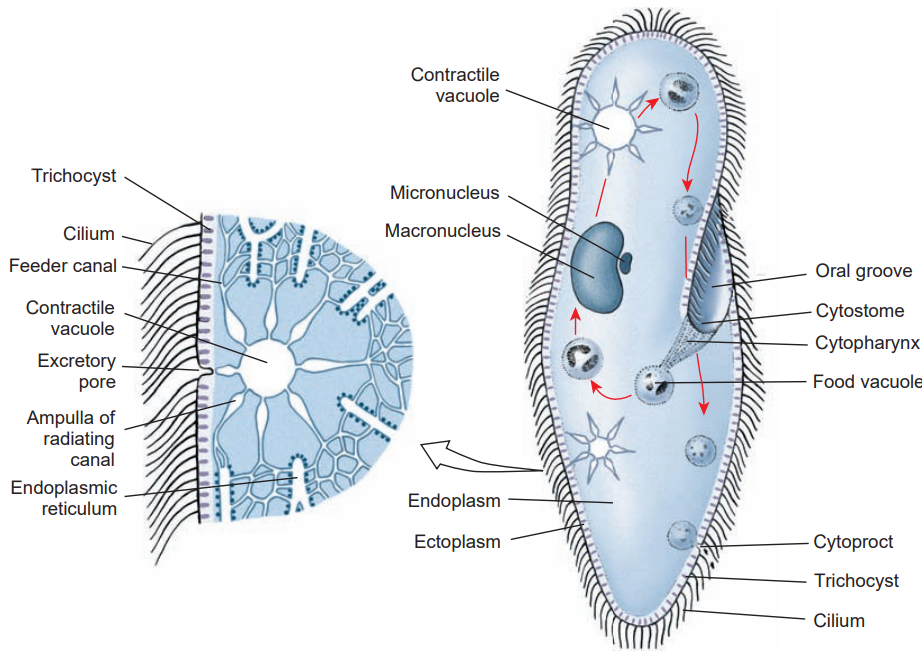
Saprozoic feeding may be by pinocytosis or by transport of solutes directly across the outer cell membrane. Direct transport across a membrane may be by diffusion, facilitated transport, or active transport. Diffusion is probably of little or no importance in nutrition of protozoa, except possibly in some endosymbiotic species. Some important food molecules, such as glucose and amino acids, may be brought into a cell by facilitated diffusion and active transport.
Useful External Links
- Protozoa: Structure, Classification, Growth, and Development by NIH
- The protozoan cell by Britanica
- Protozoan Cell Organelles by ScienceDirect