
The Origin of Respiratory Gills
In ascidian and doliolid tunicates, and in amphioxus, the gills are ciliated food collecting devices, trapping particles on mucous nets produced by the endostyle; blood flowing through them is probably de-oxygenated since the ciliary tracts of the gill bars must use more oxygen than is provided by the water flowing through them. The respiratory gills of larger and more complex chordates are likely to have been derived from a filtering arrangement of the kind seen in amphioxus, the significant step being the change from ciliary to muscular movement of water through the gill. This change, which led to the arrangement seen today in the lamprey ammocoete larva, presumably came about as a consequence of the demand for a higher filtering rate than cilia alone could provide.
When this more efficient filtering system allowed increase in body size beyond that where simple diffusion across epithelial surfaces sufficed for gas exchange, respiratory gills became specialized. In lampreys, a significant proportion of the oxygen needed is still gained across the skin, despite the development of respiratory gills. Cutaneous respiration is the only source of oxygen for many larval fishes, and is important for some adult teleosts, for example, the Antarctic icefishes lacking hemoglobin.
Respiration of Fish Larvae
Elasmobranchs, and teleosts with large eggs, such as salmon, hatch with functional gills, a well-developed circulatory system, and blood cells containing hemoglobin. Most teleosts, however, hatch as much smaller larvae depending on cutaneous respiration across the body surface. Since many of these small transparent larvae live a pelagic existence, where oxygen is plentiful, cutaneous respiration suffices, so hemoglobin is not needed and might make them conspicuous to predators. The leptocephalus larvae of elopomorph teleosts, for example, may be quite surprisingly large (some notoacanth leptocephali are 2 m long) but all are laterally compressed so that diffusion distances are small), and, lacking hemoglobin, are exceptionally transparent.
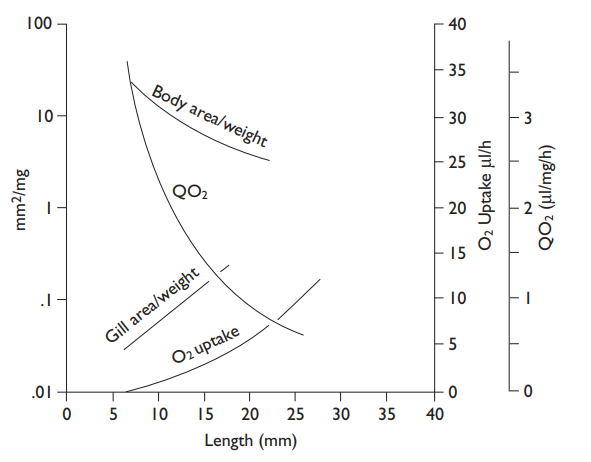
As larvae increase in size, two changes take place that affect respiration profoundly: the surface-to-volume ratio becomes smaller, so that the surface for cutaneous respiration becomes relatively smaller, and the pathways for the diffusion of gases and metabolites become longer. A size is reached when gill respiration (vastly increasing the area for diffusion) becomes essential, especially in very active fishes. The development of hemoglobin about the same time, or a little later, increases the oxygen-carrying capacity of the blood. The chemical form of hemoglobin then changes with age, as judged by electrophoretic banding patterns.
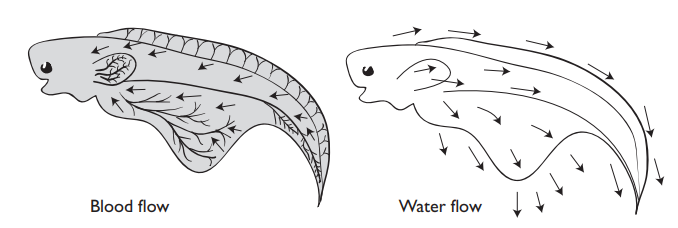
Larvae living in hypoxic environments may generate convective water flow along the body, for example in the lungfish Neoceratodus forsteri by means of cilia or by movement of the pectoral fins in Monopterus albus. In Atlantic salmon alevins, pectoral fin movements seem to draw water over the gills. Alternatively, they may undertake regular short bouts of swimming to move them out of the water they have de-oxygenated.