
The ancient fish ancestors were presumably small and fusiform, looking somewhat like amphioxus (Branchiostoma), and many modern fish have retained this streamlined kind of shape, built around an axial notochord, more or less reduced in most by the development of vertebral elements around it. But every other kind of body shape has evolved: globular (puffer fish and some angler fish); elongate (eels, pipefish, ribbon fish, agnathans); compressed dorsoventrally (rays); and laterally (sunfish, flatfish, many coral reef fish). Perhaps the most extreme body shapes are those of tropical seahorses, such as the leafy sea dragon (Phycodurus) which looks like animated seaweed, and the many curiously shaped larvae which are quite different to the adults. Living Fishes vary so much that they are hard to categorize and define briefly. Indeed, special geometric methods (thin plate spines) are being developed to compare fish body shapes (Parsons et al., 2003).
This extraordinarily wide range of body shape is perhaps best appreciated when diving off coral reefs, where Gerstner (1999) showed how body and fin variations affected maneuverability and behavior, but similar diversity is seen in the deep sea and in large tropical rivers. Whatever shape they are, most fish have a series of unpaired fins, and paired pectorals and pelvics. Paired fins are absent in lampreys and hagfish, and were thought to have arisen in gnathostome ancestry by the division of a continuous lateral fin-fold. This famous hypothesis, published by the distinguished Cambridge embryologist F. M. Balfour (1878) the year before he perished climbing in the Alps, held sway for over a century. But it is now realized that the outgrowths from the lateral plate and body wall giving rise to the unpaired fins are determined by HOX gene expression boundaries, and pectoral (the earliest) and pelvic fins are not parts of an extensive fin-fold either in ontogeny or phylogeny.
Paired fins confer the possibility for delicate maneuverability, and, like body shape, vary greatly in shape and structure. Shark fins are stiffened at their bases by cartilaginous plates bearing radial cartilages, and from the radial cartilages bristle-like collagenous rays (ceratotrichia) pass to the edges of the fins. Unfortunately, when boiled, this kind of fin provides a gelatinous soup held by some to be a delicacy, and the demand for shark fins has led to increased fishing effort, and serious decreases in shark populations. This kind of fin cannot be furled, but muscles attached to the basal cartilages can tilt it. The radials of the pectorals and pelvics are much elongated in rays, so that the (rather complicated) muscles inserting on them flex the pectorals up and down as the ray wings its way along.
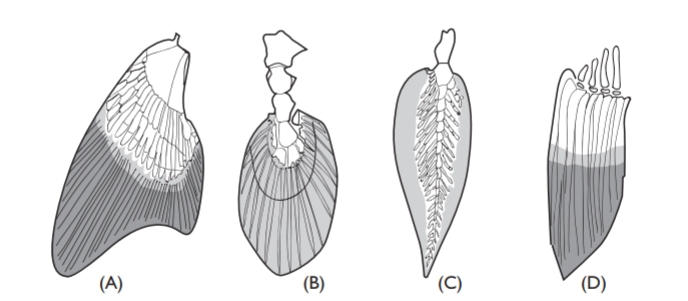
But it is teleosts that have much the most flexible and versatile fin muscula-ture. Flexible jointed rays (lepidotrichia) have basal muscles which enable these superbly designed fins to be folded and make the most delicate propul-sive movements. The precision of such movements is astonishing, best seen perhaps in seahorses and knifefishes. Sometimes, as in sea robins and gurnards (Triglidae), free pectoral fin rays are used to sample and “taste” the bottom as the fish walks forwards. However, not all teleosts have flexible fins. In the pectoral fins of marlins (Makaira), the fin rays are ossified together to form a very stiff foil for maneuvering, and the foil needs to be stiff and strongly built because marlins are capable of high speeds and the stresses in maneuvering are correspondingly high. If not in use, marlin pectorals (unlike those of sharks) can be folded flat against the body.
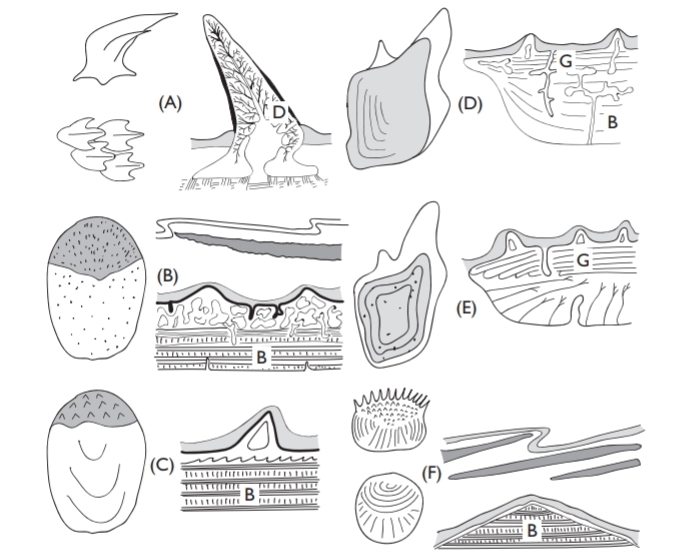
The body is usually covered with more or less conspicuous overlapping scales or denticles, overlain by an outer epithelium. Sometimes, as in eels, the scales are buried under thick epithelial layers and even when the body is armored by thick scales (Latimeria), or tough denticles (sharks and many acanthoptery-gians), the scales are still covered by an epithelial layer. In herring, and probably in other fish, this outer layer is impermeable, and, if damaged, osmotic forces cause the scale to swell. Scale shape, ornamentation, and structure vary in different fish groups, and is important in the classification of fossil fishes, as Agassiz first showed. On shape, structure, and composition, scales are now classed as they were by Agassiz: placoid, ganoid, cosmoid, ctenoid, and cycloid, although the last two really have basically the same structure.
A difficulty with the first three scale types, is that structure is not always well known, and it is not always clear whether the different variations in, say, “enamel” or enameloid tissue are significant. Studies of scale structure have undergone something of a renaissance recently, and, although scarcely a hot topic, it is worth checking older texts against recent work. Smith and Hall (1990) have reviewed the origins in phylogeny and ontogeny of different skele-tal and scale materials, concluding that dentine was the most primitive and that both the cranial endoskeleton and the exoskeleton were derived from neural crest material in the early Ordovician fishes.
Placoid scales characterize elasmobranchs (they are often termed dermal denticles) and have a vascular core of dentine capped by a thin acellular layer of “enamel,” rather similar to our own teeth. This is the only type of scale that does not increase in size as the fish grows, instead other scales are intercalated. Cosmoid scales of fossil lungfish and crossopterygians are only seen much modified in living fish, for present-day lungfish and Latimeria have lost the original outer cosmine dentine-like layer and are built of two bony layers with a thin covering of “enamel”. Ganoid scales, characteristic of Polypterus, sturgeons, and paddlefish (Polyodon) are made up of an outer acellular ganoin layer, above layers of bone. Although the scales on gars (Lepisosteus) look externally rather like those of Polypterus, the basal bony plate has a different canal arrangement. Teleosts usually have either cycloid or ctenoid scales, the latter perhaps having a hydrodynamic func-tion, as may also have shark denticle grooving. The scales of Polypterus have an unusual role in breathing.
Apart from the elasmobranch placoid scales or denticles, all other kinds grow as the fish does, and so can be used to age the fish, especially in temperate waters, where growth is seasonal. Elasmobranchs are not easy to age, but growth rings (none too easy to calibrate in years post partum) can usually be seen in vertebral centra stained in various ways, or in fin spines.