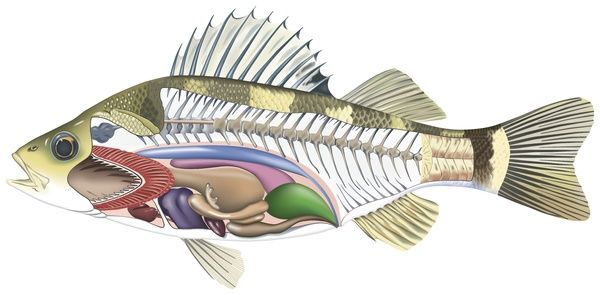
Apart from a brief foray into the Agnathan kidney, we have so far considered almost entirely extrarenal routes of ion balance and excretion, and the role of the kidney has hardly been mentioned. The elongate gnathostome fish kidney is mesonephric, and like those of adult lampreys, retains a segmental struc-ture. This is most obvious in their blood supply, which is essentially venous with a renal portal blood supply, and they operate at low systemic blood pressures not above 2.67 kPa (20 mm Hg). Fish kidneys are particularly interesting because the nephrons in different fishes are remarkably diverse and this makes it possible to infer (even without micropuncture), the func-tions of the different segments by comparing the nephrons of fish living in dif-ferent habitats .
The most striking modification of the nephron is found in some marine fish such as the angler Lophius where the glomerulus is much reduced or lost com-pletely. So far, some 30 species of marine teleosts in six families have been found to have aglomerular nephrons, and there are even some aglomerular Siamese syngnathids which have secondarily entered freshwater! As so often in physiology, the study of special cases has been rewarding, and the aglomerular kidney is no exception, for it was here that tubular secretion was first demonstrated in 1928 by the excretion of phenol red in Lophius and inulin was found (paradoxically!) to be the tracer of choice for glomerular filtration rate (GFR). The enormously large “giant” cottid nephrons are ideally suited for micropuncture studies, but as yet (so far as the authors are aware), remain to be investigated.
There are wide simultaneous variations in GFR and urine production in normal fishes, without change in urine osmolarity, and it seems that these involve recruitment of glomeruli under varying conditions. Since systemic blood pressure is always low, even small variations in pressure can “shut down” glomeruli or bring them into use, and coupled with linked changes in tubular absorption, can greatly change urine production without change in urine osmolarity. In the trout, for example, GFR measurements in single nephrons have shown that in freshwater, around 45% of the nephrons are filtering, whereas in seawater only 5% filter.
This is a flexible system, well suited to the mainly automatic regulation of urine production, particularly so for euryha- line fishes suffering changing osmotic loads as they move to waters of differ-ent salinity. Recent work on perfused trout and also perfused dogfish kidneys have shown that, as in mammals, in addition to the systemic renin-angiotensin (RAS) system, there are local systems producing angiotensin II. Addition to the perfusate of the inhibitor captopril (which blocks the angiotensin converting enzyme), leads to profound glomerular diuresis, and to changes in salt excretion, indicating an RAS system within the kidney itself.
As we should expect, the changes in GFR leading to changes in urine pro-duction are also under the control of pituitary hormones such as prolactin, isotocin, and arginine vasotocin: hormonal release itself being controlled by acentral nervous system osmoreceptor.
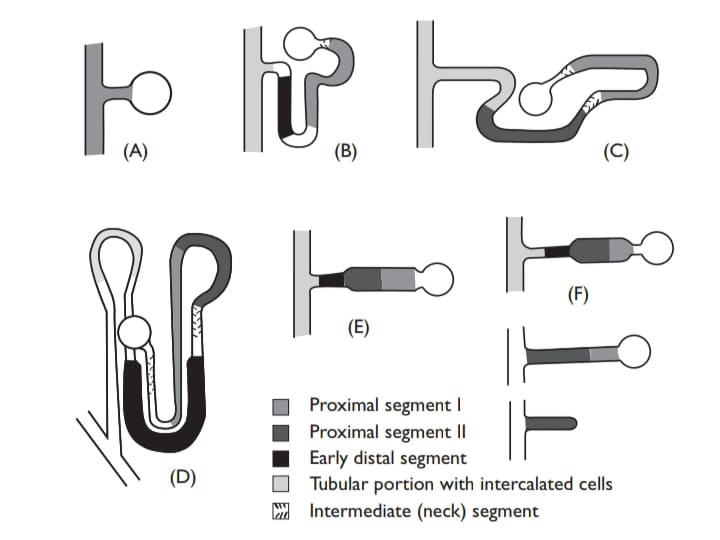
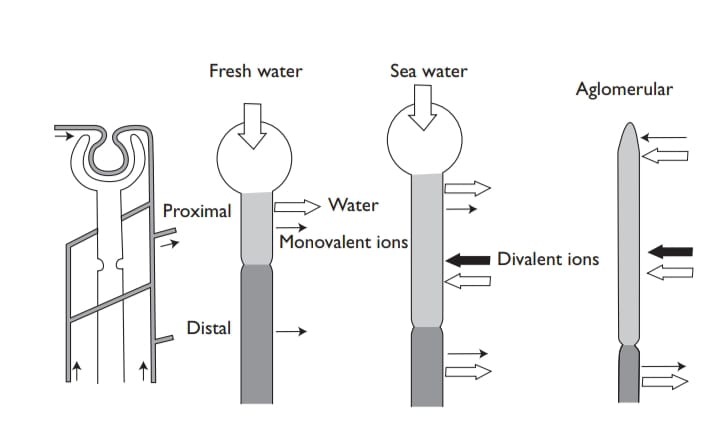
Tubular structure and arrangement varies much in different fishes . Apart from the hagfish, much the simplest is that of the teleost (hardly surprising recollecting the importance of extrarenal routes that were con-sidered a few pages back). In freshwater fishes, the proximal segment absorbs water and some monovalent ions, and the distal segment absorbs monovalent ions. In marine fishes, where the glomeruli are often much reduced, urine flow is significantly greater than GFR. The proximal tubule secretes Cl– and Na+(driven by water excretion), but it also secretes divalent ions, in particular Mg2+ and SO42+, which enter the plasma via the water the fish drinks. Both ions are at much higher levels in the sea (around 50 mM Mg2+ and 25 mM SO42+) than in the plasma. These ions are not excreted extrarenally, and although it would scarcely do the marine teleost nephron justice to consider it purely as amagnesium sulfate pump, this is certainly one of its main roles.
The distal seg-ment (lacking in many acanthopterygian) mainly absorbs water, and although some monovalent ions are excreted, extrarenal routes are more important for these. After the distal region, collection ducts of various sizes in different species pass to the urinary bladder, where (since it is well-adapted for electrophysiology) various transport mechanisms have been figured out. In marine fish, an amiloride-sensitive electrogenic component and coupled mechanism are involved in NaCl uptake, and the bladder is permeable to water where it is resorbed. In freshwater, bladder water permeability in the starry flounder (Platichthys stellatus) is six times less than in seawater so that less water is resorbed in freshwater than in seawater.