
In all fish, the body is built around an axial notochord, still present in adult agnathans, sturgeons, and Latimeria, but more or less reduced in most adult fish by the development of vertebral elements around it. This provides the incompressible strut flexed by the serial myotomal muscle blocks, and pro-tects the spinal cord, while anteriorly, some kind of cranium protects the brain, in all basically similar in structure. All fishes also share gills for gas exchange, a two-chambered heart, and a circulatory system of essentially the same plan, and a similar complement of viscera.
There are, however, striking differences between the internal structure of hagfish and lampreys, and all other fishes. For instance, in the two agnathans, the branchial skeleton is attached to the cranium and lies outside the gill pouches (in jawed fishes it is free from the cranium and supports the gills directly); there is only a single nasal opening leading to a nasohypophyseal sac; “lymphatic” vessels are lacking, and all nerve fibers are non-myelinated.
But of course, much the most significant difference is the development of hinged jaws in the gnathostome fish, accounting for their overwhelming success compared with the very few living survivors of the agnathan radiations between the Ordovician and Devonian. Once jaws had appeared, a whole series of changes from the agnathan level of organization became possible. The appearance of jaws came together with a more streamlined body and paired fins to permit the accurate control of the movements needed to snap up larger and faster prey, and a larger brain to control these movements. New gnathostome fishes of several kinds soon became dominant.
The two main gnathostome groups, the sharks, rays, and chimaeras (elasmo-branchiomorphs), and the different kinds of bony fishes, are less different than agnathans and gnathostomes but have a very distinct internal structure and biochemistry. Most obviously, the elasmobranchiomorph skeleton is almost entirely cartilaginous. The adult skull remains as a cartilaginous neurocranium to which are fused capsules for olfactory organs and the ear. This results in a curious-looking braincase (especially in Holocephali), to which the palato- quadrate and Meckel’s cartilages of the upper and lower jaw are attached. The vertebral column is rather simple; circular biconcave centra are separated by intervertebral discs and bear neural arches dorsally. Ventrally, there are either short ribs or (in the tail region) hemal arches.
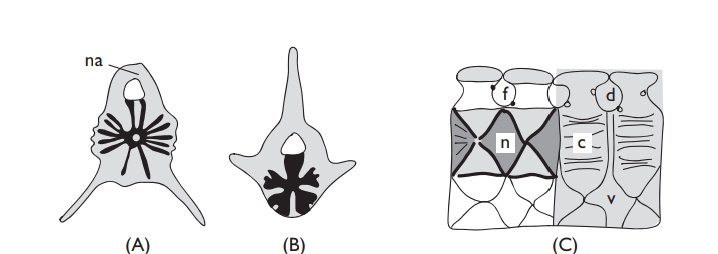
Such a relatively straightforward axial skeleton contrasts with the more complex skull and vertebral column of bony fishes. Although the skull begins in development as a cartilaginous neurocranium similar to that in sharks, it is soon altered by endochondrial bones ossifying in this cartilaginous framework, and by the addition of many dermal (membrane) bones, supposedly derived from a scale layer in the skin. The end result is a complicated compos-ite structure and that the homologies of the different bones are by no means easy to sort out. Thus, for example, the large paired dermal bones above the teleost orbit, which overlie the pineal, were long known as frontals but are now accepted as the equivalent of tetrapod parietals. Gregory (1959) gives beautiful illustrations of teleost skulls, but watch for nomenclatural problems.
In adult bony fish, the jaws are also a mix of endochondrial and membrane bones: separate ossifications in the palatoquadrate cartilage produce the pala-tine, pterygoid, mesopterygoid, and quadrate (some with dermal contribu-tions), and these are covered at the edges of the upper jaw by the dermal membrane premaxilla, maxilla, and jugal. The dermal dentary makes up most of the lower jaw, for Meckel’s cartilage only ossifies at its hinder end to form the articular. The vertebral column is normally similar to that in elasmobran-chiomorpha, with biconcave centra (although with more complex ribs).
The major differences in the viscera are the presence of a gas-filled swim-bladder in most bony fishes, although lost in some, and in others, such as myc-tophids and Latimeria, filled with lipid, and in the reproductive system. Elasmobranchiomorphs always have internal fertilization, sometimes accompanied by bizarre ways of nourishing embryos in the uterus, this is known in some bony fish but is uncommon. The spiral valve of the elas-mobranchiomorph gut is only found in a few bony fish (Amia, Polypterus, and osteoglossids) where it is less complex. Finally, bony fish lack the elasmobran-chiomorph salt-secreting rectal gland and high urea content but, curi-ously, Latimeria has both.