
A typical hydroid has a base, a stalk, and one or more terminal zooids. The base by which colonial hydroids attach to the substratum is a rootlike stolon, or hydrorhiza, which gives rise to one or more stalks called hydrocauli. The living cellular part of the hydrocaulus is a tubular coenosarc, composed of the three typical cnidarian layers surrounding the coelenteron (gastrovascular cavity). The protective covering of the hydrocaulus is a nonliving chitinous sheath, or perisarc. Attached to the hydrocaulus are individual polyp animals, or zooids. Most zooids are feeding polyps called hydranths, or gastrozooids.
They may be tubular, bottle-shaped, or vaselike, but all have a terminal mouth and a circle of tentacles. In thecate forms, such as Obelia, the perisarc continues as a protective cup around the polyp into which it can withdraw for protection. In others the polyp is (athecate) naked. In some forms the perisarc is an inconspicuous, thin film. Hydranths capture and ingest prey, such as tiny crustaceans, worms, and larvae, thus providing nutrition for the entire colony.
After partial extracellular digestion in a hydranth, the digestive broth passes along the common gastrovascular cavity where it is absorbed by gastrodermal cells, and intracellular digestion occurs. Circulation within the gastrovascular cavity is a function of the ciliated gastrodermis but is also aided by rhythmical contractions and pulsations of the body.
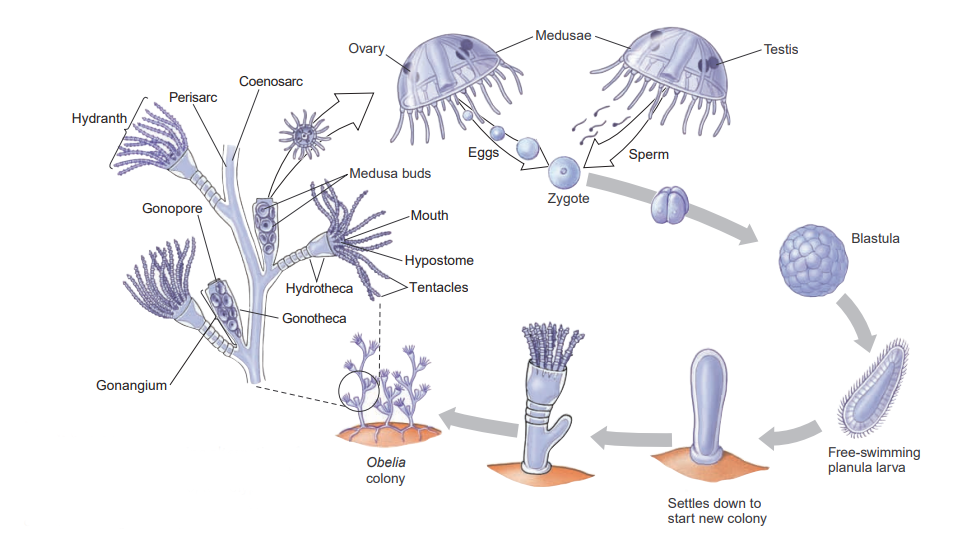
Obelia is a thecate hydroid, its polyps as well as its stems being protected by continuations of the nonliving covering.
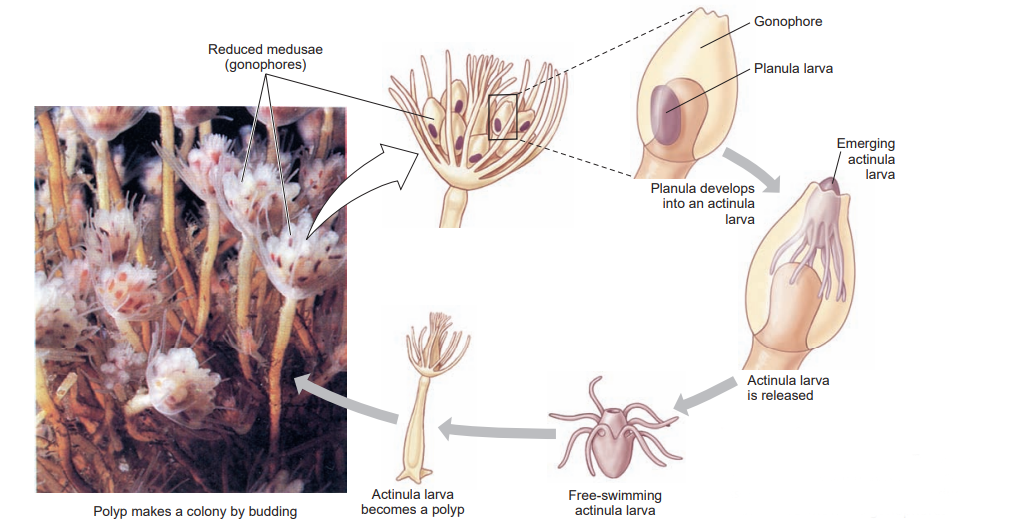
Colonial hydroids bud off new individuals, thus increasing the size of the colony. New feeding polyps arise by budding, and medusa buds also arise on the colony. In Obelia these medusae bud from a reproductive polyp called a gonangium. Young medusae leave the colony as free-swimming individuals that mature and produce gametes (eggs and sperm). In some species medusae remain attached to the colony and shed their gametes there. In other species medusae never develop and gametes are shed by male and female gonophores.
Embryonation of the zygote produces a ciliated planula larva that swims freely for a time. Then it attaches to a substratum to develop into a minute polyp that gives rise, by asexual budding, to the hydroid colony, thus completing the life cycle. Hydroid medusae are usually smaller than scyphozoan medusae, ranging from 2 to 3 mm to several centimeters in diameter. The margin of the bell projects inward as a shelflike velum, which partly closes the open side of the bell and is used in swimming. Muscular pulsations that alternately fill and empty the bell propel the animal forward, aboral side first, with a weak “jet propulsion.” Tentacles attached to the bell margin are rich in nematocysts.
The mouth opening at the end of a suspended manubrium leads to a stomach and four radial canals that connect with a ring canal around the margin. This ring canal connects with the hollow tentacles. Thus the gastrovascular cavity is continuous from mouth to tentacles, and gastrodermis lines the entire system. Nutrition is similar to that of hydranths.
The nerve net is usually concentrated into two nerve rings at the base of the velum. The bell margin has a liberal supply of sensory cells. It usually also bears two kinds of specialized sense organs: statocysts, which are small organs of equilibrium, and ocelli, which are light-sensitive organs.
The roles played by ectoderm and endoderm during the formation of hydromedusae have been investigated in one species ( Podocoryne carnea). Here, as is typical for a hydrozoan, medusa buds are produced on the sides of gonangia by lateral budding. The buds have three cell layers: ectoderm, endoderm, and a unique derivative of ectoderm called the entocodon. Portions of the entocodon differentiate into smooth and striated muscles. Further smooth muscles in the velum and tentacles originate from ectoderm. The reader may recall that cnidarians lack true mesodermally derived muscle, using epitheliomusclular cells for contraction of polyps and of nonhydrozoan medusae. Thus the presence of smooth and striated muscles in hydrozoan medusae is surprising, as is the ectodermal origin of these muscles.
Freshwater Medusae
The freshwater medusa Craspedacusta sowberii (order Hydroida) may have evolved from marine ancestors in the Yangtze River of China. Probably introduced with shipments of aquatic plants, this interesting form has now been found in many parts of Europe, throughout the United States, and in parts of Canada. Medusae may attain a diameter of 20 mm.
The polyp phase of this animal is tiny (2 mm) and has a very simple form with no perisarc and no tentacles. It occurs in colonies of a few polyps. For a long time its relation to the medusa was not recognized, and thus the polyp was given a name of its own, Microhydra ryderi. On the basis of its relationship to the jellyfish and the law of priority, both polyp and medusa should be called Craspedacusta (N.L. craspedon, velum, +Gr. kystis, bladder).
Useful External Links
- Obelia by Vedantu
- Obelia – Structure, Diagram, Life Cycle by Toppr
- Obelia Hydroid by Florida State University